C’est sans doute très péniblement que sortirent de l’onde pour s’avancer sur la terre ferme d’aventureux poissons dressés sur ce qui n’était jusque-là que les rayons de leurs nageoires. Cela s’est passé voici plus de 350 millions d’années, au Dévonien. Mais ce n’est que depuis peu que quelques esprits curieux ont l’esprit taraudé par cette question existentielle, bien normale eu égard à leur statut de bipède dominant : les rayons des nageoires des poissons d’hier sont-ils les doigts des mains et pieds des tétrapodes qui galopent aujourd’hui ? En sont-ils le prolongement ? Les homologues ?
Si on le titille, le paléontologue ordinaire (Homo sapiens prefossilis), répond preuves fossiles à l’appui que c’est peu à peu que se sont transformées les extrémités des nageoires en digitales empreintes. On a d’abord disent-ils marché au fond de l’eau, au Dévonien, sur la pointe des rayons, avant de gagner peu à peu d’abord une plage, si possible ensoleillée, puis la terre ferme pour s’y gaver d’herbages ou de gras insectes qui déjà l’habitaient. Pour preuve, s’il est en verve, il dressera le portrait de l’ancêtre Tiktaalik roseae, ce « grand poisson des basses eaux dévoniennes très âgé » (375 ma), mais toujours très vif dans la mémoire collective des paléontologues, et croqué avec talent et précision par Neil Shubin (1). Il ressemble beaucoup au coelacanthe d’aujourd’hui planqué dans les eaux comoriennes, dont les ancêtres ont pu croiser Tiktaalik, et connu bien d’autres aventures, jusqu’à se trouver ballotés dans des nappes alpines comme rapporté dans notre DinOblog par Lionel Cavin (ici)
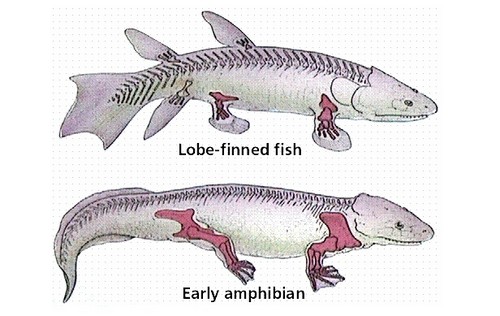
Et si on comparait les extrémités des rayons des nageoires d’un coelacanthe avec les doigts d’un tétrapode ?
Cela ne convainc qu’à demi le généticien commun (Homo sapiens caecus), et on peut le comprendre, surtout s’il est de la tribu dite des « EvoDevos ». Attention cette communauté ne cultive pas une religion sectaire comme le laisserait penser l’acronyme prononcé à la hâte : les EvoDevos s’occupent de génétique du développement, et ce pour mettre en lumière quels gènes autorisent ou interdisent les constructions qui font s’édifier les organes d’un embryon pour au final fabriquer un joli adulte très présentable.
On appelle les objets de leur attention des gènes architectes, et ils appartiennent à une quadruple fratrie, digne des frères Aymon : les Hox A,B,C,D, qui chevauchent autant de montures chromosomiques, chacun la sienne, pas d’échange possible. Plus jardiniers que guerriers, les principaux outils des frères Hox sont une batterie de lances d’arrosage, déclencheurs pour les cellules germinales qu’ils aspergent et dont ils commandent la croissance : eau chaude, les méristèmes multiplient les cellules, eau froide, arrêt du processus. Et souvent, il faut même de l’eau mitigée : 2 ou 3 Hox doivent être associés. Dans le sabir EvoDevo, ce sont des clusters.
Pour fabriquer les extrémités distales des os, les Hox activent ou non la formation de ces terminaisons appelées à soutenir les différentes parties du squelette. Leur rôle est donc essentiel et c’est leur chimie qui commande la construction, l’architecture des corps, en particulier l’ossature du squelette. Mais attention chaque Hox dispose de tout un jeu de robinets distribués de l’avant à l’arrière. Chacun a une tâche spécifique et pour les identifier et suivre leur activité, les généticiens les ont étiquetés et numérotés. Dans le cas qui nous occupe qui est de fabriquer des doigts, le cluster incriminé est Hox 3A assisté par Hox 5D.
Si les doigts des tétrapodes sont vraiment homologues des rayons des nageoires d’antan, il doit être possible de trouver chez les poissons d’aujourd’hui les mêmes capacités de différenciation génétique, les mêmes Hox, qui permettent à leur rayons de pousser et aux doigts des tétrapodes de se différencier. Et l’on étudiera de près le développement des rayons de nageoire de l’actuel poisson zèbre comparé à celui des membres et doigts de la souris.
Il n’échappera à personne qu’une nageoire de poisson est différente dans sa construction d’un membre de tétrapode : elle est faite d’une pièce, alors que le membre d’un tétrapode est construit de plusieurs éléments, soit d’un bras et d’une main avec des doigts séparés par un poignet, soit d’une jambe et d’un pied avec autant de doigts séparés par les os de la cheville.
Il fut d’abord constaté que l’action des gènes HoxD se faisait en deux temps chez les tétrapodes comme ici illustré.
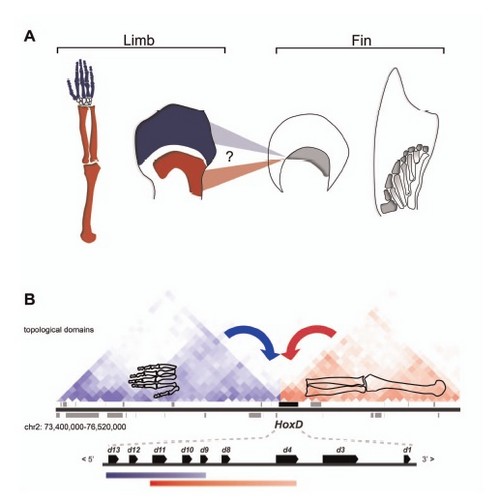
Les mécanismes de régulation et la question de l’homologie « rayon des nageoires des poissons » = « doigt des tétrapodes ». Le mode d’action et d’expression des gènes sur les extrémités des os des membres des tétrapodes sont loin d’être parfaitement compris. Le bras d’un humain (à gauche) est sous l’influence du gène HoxD qui en deux phases d’activation (bleue du côté distal et rouge du côté proximal) vont induire son développement et sa croissance. Au même stade de développement, les rayons des nageoires des poissons ne subissent qu’une seule phase d’activation. Cette zone de croissance chez le poisson est-elle l’exact homologue de la zone distale ou de la zone proximale telle qu’elles sont identifiées chez les tétrapodes ? A-t-elle des potentialités bimodales de développement ?
L’Hox5D a été interprété comme à la fois l’agent qui influe sur le développement des extrémités distales des tétrapodes et aussi sur la zone proximale.
Le problème est de mettre en évidence si cette influence bimodale préexiste déjà chez les poissons. S’il en est ainsi, d’évidence les doigts des tétrapodes seraient les exacts homologues de l’extrémité des rayons des poissons. D’après Fig.1 in référence 2.
Pour déceler si la capacité à fabriquer des doigts est déjà présente chez les ancêtres poissons, et donc montrer l’homologie d’un tel caractère, il faut se demander si le cluster HoxA a les mêmes caractéristiques que son confrère HoxD. Si c’est le cas, cela suggèrerait que les doigts sont bien homologues des rayons des nageoires et sont construits par les mêmes mécanismes de croissance. On aurait ainsi la preuve que les capacités de régulation qui permettent aux doigts de se développer existaient avant que des poissons ne se lancent hors de l’eau pour gagner la terre ferme.
Et on vient de découvrir que la réponse à cette question d’apparence simple, prête à discussions et interrogations (2). D’évidence les Hox3A et Hox5D impliqués d’une part chez le poisson zèbre, d’autre part chez la souris n’ont pas tout à fait le même domaine d’influence comme illustré sur la figure 3.
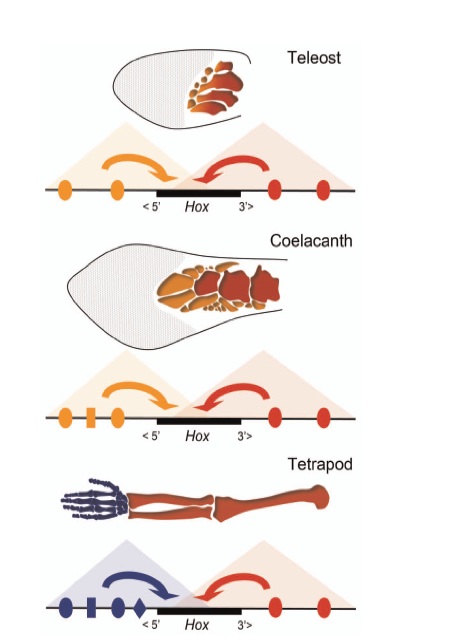
Champ d’influence des Hox3A et Hox5D chez la nageoire du poisson (Téléostéen et coelacanthe) et le membre du tétrapode (bras humain). Eu égard à leur position dans le gène, ils ont un domaine régulateur défini. Celui-ci est ici figuré par des triangles de couleur différente et des flèches de couleur correspondante indiquent leurs objectifs mérismatiques. Chez le poisson, ce mécanisme peut modeler la partie proximale (en rouge) jusqu’à celle distale (en orange). Chez les tétrapodes, ici un bras humain, il y a eu acquisition d’un nouvel accélérateur (en bleu) qui a modifié le mode d’action dans la région distale. D’après Fig. 7 in référence 2.
Bien que les mécanismes qui règlent l’expression de HoxA et HoxD soient comparables chez l’un et l’autre, quand les régions génomiques qui règlent l’expression de gène Hox dans des nageoires de poisson sont injectés à des souris transgéniques, les gènes en question déclenchent la prolifération de cellules osseuses. Mais seulement le segment de membre proximal (le plus proche le corps) est affecté, alors que la région distale et donc les doigts ne subissent aucune transformation.
Il faut conclure que si les poissons ont les robinets Hox A et D, et donc l’eau froide à tous les étages, leurs successeurs tétrapodes bénéficient eux du confort moderne : ils peuvent dégourdir leurs doigts à l’eau mitigée. Dès lors on ne peut pas considérer que l’extrémité des rayons des nageoires est l’exacte homologue des doigts des tétrapodes. Ces derniers peuvent se développer grâce à une innovation qui n’est pas inscrite dans le bail plus qu’emphytéotique légué par leurs ancêtres poissons. Non, les rayons des nageoires ne sont pas les homologues sensu stricto des doigts des tétrapodes.
Références :
N. Shubin. 2008. Your Inner Fish: A Journey into the 3.5-Billion-Year History of the Human Body. New York: Pantheon Books.
Woltering JM, Noordermeer D, Leleu M, Duboule D (2014). Conservation and Divergence of Regulatory Strategies at Hox Loci and the Origin of Tetrapod Digits. PLoS Biol 12(1): e1001773. doi:10.1371/journal.pbio..
 Tags :
Tags : Publié dans : Evolution,Nouveautés
Les commentaires et les pings ne sont pas autorisés.






[...] C’est sans doute très péniblement que sortirent de l’onde pour s’avancer sur la terre ferme d’aventureux poissons dressés sur ce qui n’était jusque-là que les rayons de leurs nageoires. [...]
[...] Michel Fontaine est dessinateur, sculpteur et photographe d’animaux disparus surtout, mais actuels aussi. Les articles du DinOblog lui inspirent quelques dessins qui viendront désormais ponctuer les billets. Et voici ce que donne dans l’oeil de Michel la lecture de l’article de Jean-Louis Hartenberger, Contruire un doigt (ici) [...]